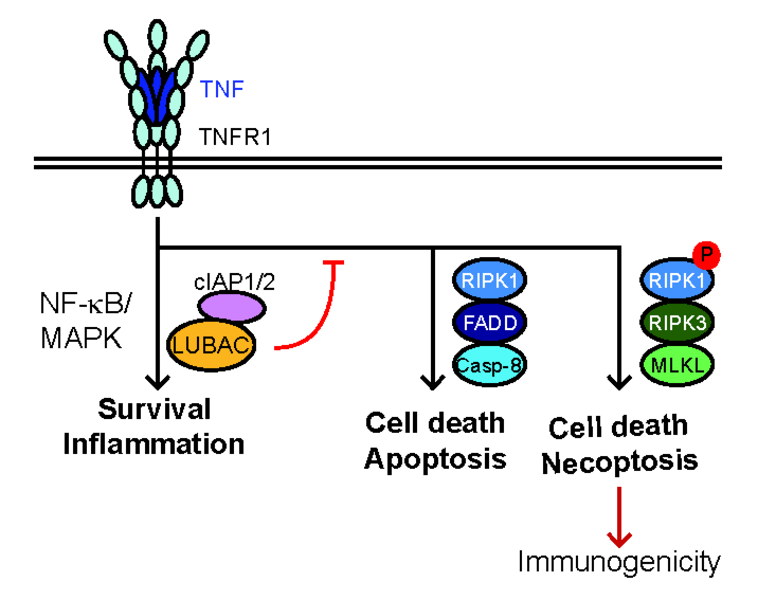
Cell death and inflammation are common features of many autoimmune and degenerative disorders. It is now clear that cell death can trigger chronic inflammation resulting in autoimmunity and degenerative diseases, including cancer. Therefore, understanding the crosstalk between death and inflammation is important to find effective therapies for such diseases. We aim to investigate the processes regulating the execution of cell death downstream of innate immune receptors. In particular, we want to uncover the importance of different modalities of cell death in autoimmune diseases and cancer, with the ultimate purpose to find to therapeutic strategies to tackle inflammation-dependent pathologies. Cell death is crucial to maintain tissue homeostasis as it is important for tissue repair and infectious processes. Consequently, many organisms can sense both damage and pathogens, a deed achieved via the so-called damage- and pathogen-associated molecular patterns (DAMPs and PAMPs, respectively) that are recognised by Pattern-recognition receptors (PRRs). The PRR family consists of Toll-like receptors (TLRs), RIG-I-like receptors (RLRs), NOD-like receptors (NLRs), and DNA sensors such as DNA-induced activator of interferon (IFN) (DAI, also known as ZBP1) and cyclic GMP-AMP synthase (cGAS). Triggering of PRRs results in the induction of inflammatory cytokines and chemokines including, among others, TNF, IL-1β and type I IFNs. These cytokines play crucial roles in triggering innate immune responses by binding to their respective receptors, including TNF receptor (TNFR) superfamily (SF) members. Binding of TNF to its cognate receptor TNFR1 results in downstream cascades that induce: i) inflammation/survival via LUBAC and cIAP1/2, by activating NF- κB/MAPK-mediated gene expression, ii) apoptosis via FADD/RIPK1/caspase-8 or, iii) necroptosis via RIPK1 autophosphorylation, RIPK3 and MLKL (Fig. 1). Recently, Caspase-8 activation by TNF was reported to induce pyroptosis via Gasdermin D and, indirectly, secondary necrosis via Gasdermin E (Fig. 1). In normal physiology, TNFR1-signalling output is skewed towards inflammation/survival; however, in autoimmune disorders or pathological conditions this balance is shifted towards cell death induction.
Figure 1
Fig. 1: TNFR1 signaling outputs.
In normal physiology, TNF induced inflammation and survival; however, in pathological conditions this balance is shifted towards cell death. Cell death can be induced by caspase-8-mediated apoptosis or RIPK3/MLKL-mediated necroptosis. Necroptosis additionally requires the kinase activity of RIPK1. Caspase-8 can further activate Gasdermin (Gsdm) D to promote pyroptosis and GsdmE, via Caspase-3, to promote secondary necrosis/pyroptosis. Necroptosis and pyroptosis are highly immunogenic due to the release of DAMPs. LUBAC and cIAP1/2 are crucial regulators of TNFR1-signaling output by preventing cell death.
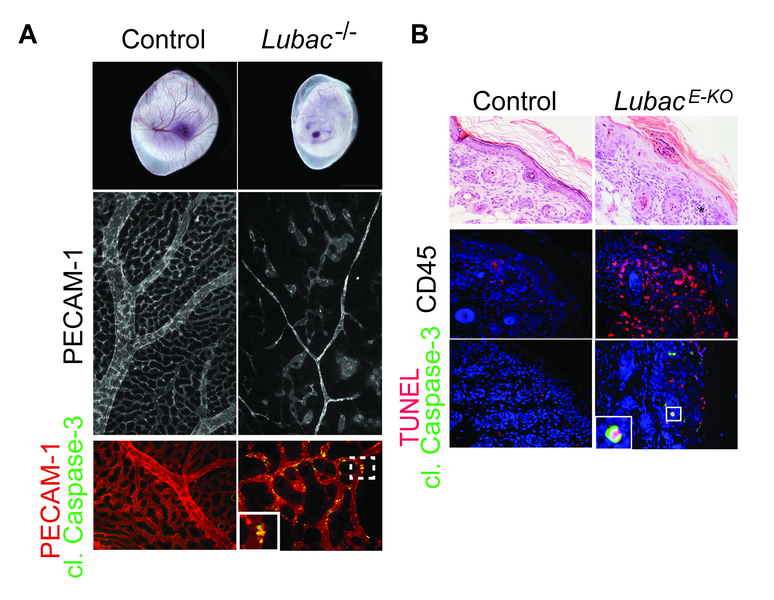
In the past years, we discovered that The Linear Ubiquitin chain Assembly Complex (LUBAC), an E3 ligase that generates linear ubiquitin chains on components of many immune receptor-signaling complexes, is crucial for survival as it prevents both apoptosis and necroptosis (Fig. 2A). In a model of LUBAC deficiency specifically in the skin, which results in severe dermatitis due to excessive cell death (Fig. 2B), we further found that combined loss of several DRs (namely TNFR1, CD95 (Fas) and TRAIL-R), but not their individual loss, prevented chronic inflammation. Thus, we identified CD95 and TRAIL-R, in addition to TNFR1, as important DRs in cell death-induced inflammation, providing support for combining DR-inhibition as treatment of inflammation-associated autoimmunity.
Figure 2
Fig. 2: LUBAC maintains tissue homeostasis by preventing cell death. A. Loss of LUBAC in the whole-body results in embryonic lethality (upper panel) due to disruption of yolk sac vasculature (upper and middle panel). Excessive cell death is detected in endothelial cells which causes the defects in vascularization (lower panel). (Peltzer, N., et al. Cell Reports 2014, Peltzer, N., et al. Nature 2018) B. Loss of LUBAC in the epidermal keratinocytes results in severe skin inflammation (upper and middle panels). Excessive cell death is observed in inflamed skin (lower panel) and is the cause of dermatitis (Taraborrelli, L., Peltzer, N., et al. Nature Communications 2018)
The general concept is that necroptosis is highly immunogenic, due to the release of immunogenic factors such as DAMPs, whilst apoptosis is immunologically silent. However, this concept has been challenged and in certain circumstances apoptosis can also trigger immunogenicity. It is not only the mode of cell death itself to dictate immunogenicity, but also the inflammatory pathways that are concomitantly activated, and the consequent release of immunogenic factors. The decision between survival and death and the cell death programs that are activated will determine a pathophysiological outcome. Therefore, it is important to define which and how cell death processes shape immune responses in health and disease.
Autoimmune Diabetes
One common autoimmune disease with a cell death signature is Type 1 Diabetes (TID), which is caused by an immune reaction against pancreatic β-cells. However, the exact mechanisms dictating β-cell demise responsible for activating immune responses during T1D are currently poorly understood. We focus on understanding the signaling cascades that mediate cell death and inflammation in T1D and in identifying β-cell death mechanisms that induce or perpetuate immunogenicity. Diabetes is also induced by obesity. Yet, in this case, it is called Type II Diabetes (TIID) and the specific immune attack of β-cells is less clear. Instead, obesity and the associated insulin resistance ultimately lead to β-cell exhaustion which reduces β-cell mass. Again, the mechanisms of β-cell demise TIID are not entirely clear. We want to study the mechanisms dictating β-cell demise in the context of autoimmunity and obesity and find common and unique features in TI/IID.
Obesity-induced inflammation
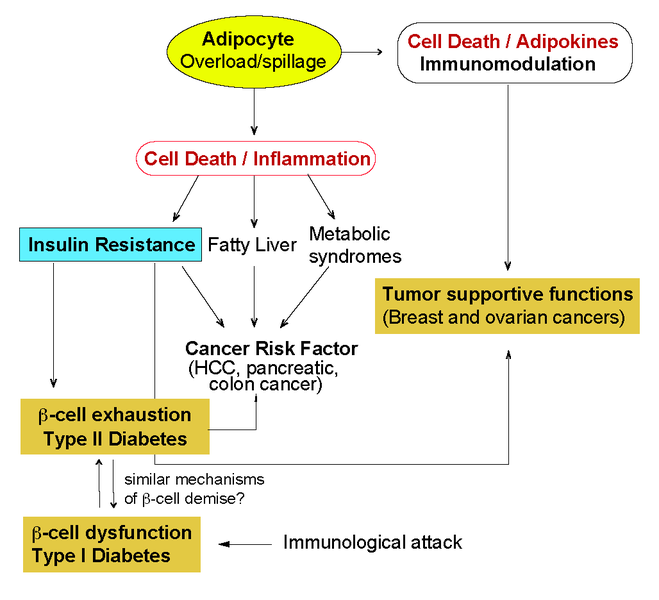
Chronic, low-grade inflammation of adipose tissue plays a key role in the pathogenesis of obesity. Both adipocytes and the macrophages that accumulate in adipose tissue during obesity are source of inflammatory cytokines and have been identified as key components in the development of chronic inflammation and consequent insulin resistance (IR) (Fig. 3). Chronic inflammation and IR ultimately lead to Type II Diabetes (TIID). Obesity-associated inflammation is a prominent cause of cancer. Increasing adiposity leads to adipocyte death and accumulation of macrophages in the so-called crown-like structures (CLS) which has been shown to have tumor supportive functions. The type of adipocyte death that dictates the formation of tumor supportive CLS as well as the molecular mechanisms leading to adipose tissue macrophages recruitment and the type of macrophages recruited are still not completely understood. We propose to study the interplay between cell death and obesity-induced inflammation and understand the contribution of adipocyte death during obesity in metabolic syndromes and tumor microenvironment.
Figure 3
Fig. 3: Obesity induced inflammation and associated complications. Scheme of the intertwined consequences of obesity-induced inflammation in metabolic diseases and cancer that are of interest for our research group.
Cell death in cancer
Gasdermins are emerging as key determinants for antitumor immunity as well as therapy response. We are currently studying how the tumor cells regulate gasdermin expression to their own advantage. Intriguingly, tumors may use gasdermin function not only to evade cell death but also to modulate the tumor microenviornemnt. We are assessing the role of gasdermins and the impact of pharmacological modulation of gasdermin-dependent functions in small cell lung cancer (SCLC), an aggressive type of solid tumor, and in chronic lymphocytic leukemia (CLL), which is highly dependent on a tumor supportive microenvironment.
Perspectives
We aim to uncover which and how cell death pathways modulate disease outcome. By building a research programme in which basic and translational research are closely aligned we strive to link fundamental cell biology to pathophysiological mechanisms to develop novel treatments options for patients.
Peltzer, N* and Annibaldi, A*. Cell Death-Related Ubiquitin Modifications in Inflammatory Syndromes: From Mice to Men. Biomedicines, doi:10.3390/biomedicines10061436 (2022) (*co-corresponding authors)
Peltzer, N.*,and Walczak, H*.Cell Death and Inflammation - A Vital but Dangerous Liaison. Trends in immunology 40, 387-402, 2019 (*co-corresponding authors)
Taraborrelli, L.*, Peltzer, N.*, Montinaro, A., Kupka, S., Rieser, E., Hartwig, T., Sarr, A., Darding, D., Draber, P., Haas, T.L., Akarca, A., Marafioti, T., Pasparakis, M., Bertin, J., Gough, P.J., Bouillet, P., Strasser, A., Leverkus, M., Silke, S., Walczak, H. LUBAC prevents lethal dermatitis by combined inhibition of TNF-, TRAIL- and CD95L-mediated cell death. Nature Communications 9: 3910, 2018 (*equal contribution).
Peltzer, N., Darding, M., Montinaro, A., Draber, P., Draberova, H., Kupka, S., Rieser, E., Fisher, A., Hutchinson, C., Taraborrelli, L., Hartwig, t., Lafont, E., Haas, T.L., Shimizu, Y., Böiers, C., Sarr, A., Rickard, J., Alvarez-Diaz, S., Ashworth, M.T., Beal, A., Enver, T., Bertin, J., Kaiser, W., Strasser, A., Silke, J., Bouillet, P., Walczak, H. LUBAC is essential for embryogenesis by preventing cell death and enabling haematopoiesis. Nature 557: 112–117, 2018.
Shimizu, Y., Peltzer, N., Sevko, A., Lafont, E., Sarr, A., Draberova, H., Walczak, H. The linear ubiquitin chain assembly complex acts as a liver tumour suppressor and inhibits hepatocyte apoptosis and hepatitis. Hepatology 65(6): 1963-1978, 2017
Peltzer, N., Rieser, E., Taraborrelli, L., Draber, P., Darding, M., Pernaute, B., Shimizu, Y., Daboh, A., Draberova, H., Montinaro, A., Martinez-Barbera, J.P., Silke, J., Rodriguez, T.A. and Walczak, H. HOIP deficiency caused embryonic lethality by aberrant TNFR1-mediated endothelial cell death. Cell Reports 9: 153-165, 2014.
Khalil, H.*, Peltzer, N.*, Walicki, J., Yang, JY., Dubuis, G., Gardiol, N., Held, W., Bigliardi, P., Marsland, B., Liaudet, L. and Widmann, C. Caspase-3 protects stressed organs against cell death. Molecular and Cellular Biology 32(22): 4523-33, 2012 (*equal contribution).
Hildebrandt X, Ibrahim M, Peltzer N. Cell death and inflammation during obesity: "Know my methods, WAT(son)". Cell Death Differ. 2022 Sep 29:1-14. doi: 10.1038/s41418-022-01062-4. Online ahead of print. PMID: 36175539
Peltzer N, and Annibaldi A (2022). Cell Death-Related Ubiquitin Modifications in Inflammatory Syndromes: From Mice to Men. Biomedicines10. doi:10.3390/biomedicines10061436.
Peltzer N (2022). Linear ubiquitin as a common regulator of cellular stress. FEBS J. doi:10.1111/febs.16427.
Cell Death and Inflammation - A Vital but Dangerous Liaison. Peltzer N, Walczak H. Trends Immunol. 2019 May;40(5):387-402. doi: 10.1016/j.it.2019.03.006. Epub 2019 Apr 16. PMID: 31003931 Review.
Endothelial Cell Killing by TAK1 Inhibition: A Novel Anti-angiogenic Strategy in Cancer Therapy. Peltzer N, Walczak H. Dev Cell. 2019 Jan 28;48(2):127-128. doi: 10.1016/j.devcel.2019.01.011. PMID: 30695692
RIPK1 and Caspase-8 Ensure Chromosome Stability Independently of Their Role in Cell Death and Inflammation. Liccardi G, Ramos Garcia L, Tenev T, Annibaldi A, Legrand AJ, Robertson D, Feltham R, Anderton H, Darding M, Peltzer N, Dannappel M, Schünke H, Fava LL, Haschka MD, Glatter T, Nesvizhskii A, Schmidt A, Harris PA, Bertin J, Gough PJ, Villunger A, Silke J, Pasparakis M, Bianchi K, Meier P. Mol Cell. 2019 Feb 7;73(3):413-428.e7. doi: 10.1016/j.molcel.2018.11.010. Epub 2018 Dec 28. PMID: 30598363
LUBAC prevents lethal dermatitis by inhibiting cell death induced by TNF, TRAIL and CD95L. Taraborrelli L, Peltzer N, Montinaro A, Kupka S, Rieser E, Hartwig T, Sarr A, Darding M, Draber P, Haas TL, Akarca A, Marafioti T, Pasparakis M, Bertin J, Gough PJ, Bouillet P, Strasser A, Leverkus M, Silke J, Walczak H. Nat Commun. 2018 Sep 25;9(1):3910. doi: 10.1038/s41467-018-06155-8. PMID: 30254289
LUBAC is essential for embryogenesis by preventing cell death and enabling haematopoiesis. Peltzer N, Darding M, Montinaro A, Draber P, Draberova H, Kupka S, Rieser E, Fisher A, Hutchinson C, Taraborrelli L, Hartwig T, Lafont E, Haas TL, Shimizu Y, Böiers C, Sarr A, Rickard J, Alvarez-Diaz S, Ashworth MT, Beal A, Enver T, Bertin J, Kaiser W, Strasser A, Silke J, Bouillet P, Walczak H. Nature. 2018 May;557(7703):112-117. doi: 10.1038/s41586-018-0064-8. Epub 2018 Apr 25. PMID: 29695863
Mitochondrial permeabilization engages NF-κB-dependent anti-tumour activity under caspase deficiency. Giampazolias E, Zunino B, Dhayade S, Bock F, Cloix C, Cao K, Roca A, Lopez J, Ichim G, Proïcs E, Rubio-Patiño C, Fort L, Yatim N, Woodham E, Orozco S, Taraborrelli L, Peltzer N, Lecis D, Machesky L, Walczak H, Albert ML, Milling S, Oberst A, Ricci JE, Ryan KM, Blyth K, Tait SWG. Nat Cell Biol. 2017 Sep;19(9):1116-1129. doi: 10.1038/ncb3596. Epub 2017 Aug 28. PMID: 28846096
The Linear ubiquitin chain assembly complex acts as a liver tumor suppressor and inhibits hepatocyte apoptosis and hepatitis. Shimizu Y, Peltzer N, Sevko A, Lafont E, Sarr A, Draberova H, Walczak H. Hepatology. 2017 Jun;65(6):1963-1978. doi: 10.1002/hep.29074. Epub 2017 Apr 10. PMID: 28120397
LUBAC deficiency perturbs TLR3 signaling to cause immunodeficiency and autoinflammation. Zinngrebe J, Rieser E, Taraborrelli L, Peltzer N, Hartwig T, Ren H, Kovács I, Endres C, Draber P, Darding M, von Karstedt S, Lemke J, Dome B, Bergmann M, Ferguson BJ, Walczak H. J Exp Med. 2016 Nov 14;213(12):2671-2689. doi: 10.1084/jem.20160041. Epub 2016 Oct 24. PMID: 27810922
Holding RIPK1 on the Ubiquitin Leash in TNFR1 Signaling. Peltzer N, Darding M, Walczak H. Trends Cell Biol. 2016 Jun;26(6):445-461. doi: 10.1016/j.tcb.2016.01.006. Epub 2016 Feb 11. PMID: 26877205 Review.
TNFR1-dependent cell death drives inflammation in Sharpin-deficient mice. Rickard JA, Anderton H, Etemadi N, Nachbur U, Darding M, Peltzer N, Lalaoui N, Lawlor KE, Vanyai H, Hall C, Bankovacki A, Gangoda L, Wong WW, Corbin J, Huang C, Mocarski ES, Murphy JM, Alexander WS, Voss AK, Vaux DL, Kaiser WJ, Walczak H, Silke J. Elife. 2014 Dec 2;3:e03464. doi: 10.7554/eLife.03464. PMID: 25443632
HOIP deficiency causes embryonic lethality by aberrant TNFR1-mediated endothelial cell death. Peltzer N, Rieser E, Taraborrelli L, Draber P, Darding M, Pernaute B, Shimizu Y, Sarr A, Draberova H, Montinaro A, Martinez-Barbera JP, Silke J, Rodriguez TA, Walczak H. Cell Rep. 2014 Oct 9;9(1):153-165. doi: 10.1016/j.celrep.2014.08.066. Epub 2014 Oct 2. PMID: 25284787
The activity of the anti-apoptotic fragment generated by the caspase-3/p120 RasGAP stress-sensing module displays strict Akt isoform specificity. Vanli G, Peltzer N, Dubuis G, Widmann C. Cell Signal. 2014 Dec;26(12):2992-7. doi: 10.1016/j.cellsig.2014.09.009. Epub 2014 Sep 22. PMID: 25246356
Fire raiser or fire accelerant? A meeting report on the 14th International TNF Conference 2013. von Karstedt S, Peltzer N. Cell Death Differ. 2014 Aug;21(8):1343-4. doi: 10.1038/cdd.2014.43. Epub 2014 Apr 11. PMID: 24722211
Ubiquitin in the immune system. Zinngrebe J, Montinaro A, Peltzer N, Walczak H. EMBO Rep. 2014 Jan;15(1):28-45. doi: 10.1002/embr.201338025. Epub 2013 Dec 27. PMID: 24375678
Role of mTOR, Bad, and Survivin in RasGAP Fragment N-Mediated Cell Protection. Peltzer N, Vanli G, Yang JY, Widmann C. PLoS One. 2013 Jun 27;8(6):e68123. doi: 10.1371/journal.pone.0068123. Print 2013. PMID: 23826368 Free PMC article.
Caspase-3 protects stressed organs against cell death. Khalil H, Peltzer N, Walicki J, Yang JY, Dubuis G, Gardiol N, Held W, Bigliardi P, Marsland B, Liaudet L, Widmann C. Mol Cell Biol. 2012 Nov;32(22):4523-33. doi: 10.1128/MCB.00774-12. Epub 2012 Sep 4. PMID: 22949508 Free PMC article.
UV-B induces cytoplasmic survivin expression in mouse epidermis. Peltzer N, Bigliardi P, Widmann C. J Dermatol Sci. 2012 Sep;67(3):196-9. doi: 10.1016/j.jdermsci.2012.05.007. Epub 2012 Jun 15. PMID: 22748311 No abstract available.
RasGAP-derived fragment N increases the resistance of beta cells towards apoptosis in NOD mice and delays the progression from mild to overt diabetes. Bulat N, Jaccard E, Peltzer N, Khalil H, Yang JY, Dubuis G, Widmann C. PLoS One. 2011;6(7):e22609. doi: 10.1371/journal.pone.0022609. Epub 2011 Jul 25. PMID: 21799917
Dr. Nieves Peltzer
Dept. of Translational Genomics & Center for Molecular Medicine Cologne - CECAD Research Center
This website uses cookies. Those have two functions: On the one hand they are providing basic functionality for this website. On the other hand they allow us to improve our content for you by saving and analyzing anonymized user data. You can redraw your consent to using these cookies at any time. Find more information regarding cookies on our Data Protection Declaration and regarding us on the Imprint.