The main interest of our laboratory is to uncover the genetic and epigenetic factors controlling the deployment of gene expression programs during vertebrate embryogenesis as well as how the disruption of these regulatory factors can lead to human congenital disease. Towards these goals, we are using a multidisciplinary approach that combines in vitro and in vivo models with biochemical, genomic and genetic engineering tools.
Enhancers are compact (~200-500bp) DNA sequences that control the activity of their target gene over long distances and in an orientation-independent manner. The importance of enhancers during embryogenesis is well illustrated by enhancer deletions or mutations in both animal models and humans that can lead to severe congenital abnormalities. Despite the develop-mental and medical relevance of enhancers, our understanding of these distal regulatory elements was rather limited until recently. The lack of a stereotypical sequence com-position, limited sequence conservation and distal position with respect to target genes, had hist-orically made the discovery of enhancers a daunting task. In the last few years, epigenomic profiling and the use of chromatin signatures have emerged as powerful and universal tools to identify enhancers.
Our laboratory previously uncovered novel chro-matin signatures that enabled not only the global identification of enhancers but also their distinction into two major classes depending on their activity: active and poised. Importantly, these chromatin signatures were shown to be universal and have been subsequently used by us and others to identify enhancers in multiple cell types and organisms. In recent years, genome-wide asso-ciation studies (GWAS) have revealed that ~90% of genetic variants associated with common human diseases occur within the vast non-coding fraction of the human genome, preferentially within enhancers.
Moreover, it has been demonstrated that changes in the regulatory activity or topology of enhancers, due to genetic variation or genomic re-arrangements, respectively, can actually lead to human disease.
We have previously described poised enhancers in embryonic stem cells (ESC) as a set of cis-regulatory elements characterized by the simultaneous presence of transcription factors, co-activators, H3K4me1 and H3K27me3. Our recent epigenomic and functional annotation of poised enhancers in mouse ESC suggested that, rather than being promiscuously used during the acquisition of somatic cell fates, poised enhancers were preferentially involved in the establishment of anterior neural identity.
Using CRISPR/Cas9 to delete poised enhancers in situ, we showed that these cis-regulatory elements are necessary for the induction of major forebrain regulators. Moreover, 4C-seq experiments revealed that poised enhancers established strong and specific physical interactions with their target genes already in the undifferentiated ESC state. Remarkably, additional 4C-seq experiments showed that such pre-existing topological conformations are polycomb dependent.
Together with additional characteri-zation of polycomb null ESC, we proposed that polycomb proteins confer poised enhancers with the appropriate regulatory topology and, thus, facilitate the timely induction of forebrain master regulators. Overall, our work illuminates a novel mechanism by which polycomb proteins might facilitate the proper establishment of developmental gene expression programs (Figure 1). This work was published in Cell Stem Cell.
The pathological consequences of structural variants disrupting 3D genome organization can be difficult to elucidate in vivo due to differences in gene dosage sensitivity between mice and humans. This is illustrated by Branchiooculofacial Syndrome (BOFS), a rare congenital disorder caused by hetero-zygous mutations within TFAP2A, a neural crest regulator for which humans, but not mice, are haploinsufficient. We recently described a BOFS patient carrying a heterozygous inversion with one breakpoint located within a Topologically Associa-ting Domain (TAD) containing enhancers essential for TFAP2A expression in human neural crest cells (hNCC). Using patient-specific hiPSCs, we showed that, although the inversion shuffles the TFAP2A hNCC enhancers with novel genes within the same TAD, this does not result in enhancer adoption or ectopic gains in gene expression. Instead, the inversion disconnects one TFAP2A allele from its cognate enhancers, leading to monoallelic and haploinsufficient TFAP2A expres-sion in patient hNCC. Therefore, our work highlights the power of hiPSCs differentiation to unveil the pathological mechanisms whereby structural variants can cause congenital abnormalities. This work was recently publihsed in Cell Stem Cell. Based on these recent findings, we are now dissecting the regulatory logic govering the compatibility and responsiveness between genes and enhancers, which we believe, includes genetic, epigenetic and structural factors.
Susceptibility to most human diseases depends on the interactions between genetic, epigenetic and environmental factors. While disease-associated genetic variants frequently occur within enhancers, their potential consequences at these genomic locations remain poorly understood. Multiple re-dundant enhancers often control key regulatory genes, which, at least in invertebrate organisms, can provide transcriptional robustness against en-vironmental stress. We postulate that due to this enhancer redundancy the pathological effects of etiological enhancer variation can only be revealed under environmental stress. Using the neural crest to model congenital disease and teratogen sensi-tivity, we will use a multidisciplinary approach that combines genomics, computational modelling and genetic engineering to systematically evaluate the importance of enhancer redundancy. By considering enhancers as hubs of gene-environ-mental inter-actions, our approach can illuminate a novel and general etiological paradigm for human disease.
2. Rada-Iglesias A#, Grosveld FG, Papantonis A#. Forces driving the three-dimensional folding of eu-karyotic genomes. Mol Syst Biol. 2018 Jun 1;14(6):e8214. PMID: 29858282.
3. Rada-Iglesias A#. Is H3K4me1 at enhancers correlative or causative. Nat Genet. 2018 Jan;50(1):4-5. PMID:29273804.
4. Cruz-Molina S, Respuela P, Tebartz C, Kolovos P, Nikolic M, Fueyo R, van Ijcken W.F.J, Grosveld F, Frommolt P, Bazzi H, Rada-Iglesias A#. PRC2 facilitates the regulatory topology required for poised enhancer function during pluripotent stem cell differ-entiation. Cell Stem Cell. 2017 May 4;20(5):689-705. PMID:28285903.
5. Nikolić M, Papantonis A, Rada-Iglesias A#. GARLIC: A Bioinformatic Toolkit for Etiologically Connecting Diseases and Cell Type-Specific Regulatory Maps. Hum Mol Genet. 2017 Feb 15;26(4):742-752. PMID:28007912.
Bleckwehl, T., and Rada-Iglesias, A. (2019). Transcriptional and epigenetic control of germline competence and specification. Curr Opin Cell Biol 61, 1-8.
Frank, S., Ahuja, G., Bartsch, D., Russ, N., Yao, W., Kuo, J.C., Derks, J.P., Akhade, V.S., Kargapolova, Y., Georgomanolis, T., Messling, J.E., Gramm, M., Brant, L., Rehimi, R., Vargas, N.E., Kuroczik, A., Yang, T.P., Sahito, R.G.A., Franzen, J., Hescheler, J., Sachinidis, A., Peifer, M., Rada-Iglesias, A., Kanduri, M., Costa, I.G., Kanduri, C., Papantonis, A., and Kurian, L. (2019). yylncT Defines a Class of Divergently Transcribed lncRNAs and Safeguards the T-mediated Mesodermal Commitment of Human PSCs. Cell Stem Cell 24, 318-27 e8.
Laugsch, M., Bartusel, M., Rehimi, R., Alirzayeva, H., Karaolidou, A., Crispatzu, G., Zentis, P., Nikolic, M., Bleckwehl, T., Kolovos, P., van Ijcken, W.F.J., Saric, T., Koehler, K., Frommolt, P., Lachlan, K., Baptista, J., and Rada-Iglesias, A. (2019). Modeling the Pathological Long-Range Regulatory Effects of Human Structural Variation with Patient-Specific hiPSCs. Cell Stem Cell 24, 736-52 e12.
Matheus, F., Rusha, E., Rehimi, R., Molitor, L., Pertek, A., Modic, M., Feederle, R., Flatley, A., Kremmer, E., Geerlof, A., Rishko, V., Rada-Iglesias, A., and Drukker, M. (2019). Pathological ASXL1 Mutations and Protein Variants Impair Neural Crest Development. Stem Cell Reports 12, 861-8.
Frank S, Ahuja G, Bartsch D, Russ N, Yao W, Kuo JC, Derks JP, Akhade VS, Kargapolova Y, Georgomanolis T, Messling JE, Gramm M, Brant L, Rehimi R, Vargas NE, Kuroczik A, Yang TP, Sahito RGA, Franzen J, Hescheler J, Sachinidis A, Peifer M, Rada-Iglesias A, Kanduri M, Costa IG, Kanduri C, Papantonis A, and Kurian L (2018). yylncT Defines a Class of Divergently Transcribed lncRNAs and Safeguards the T-mediated Mesodermal Commitment of Human PSCs. Cell Stem Cell10.1016/j.stem.2018.11.005.
Fueyo R, Iacobucci S, Pappa S, Estaras C, Lois S, Vicioso-Mantis M, Navarro C, Cruz-Molina S, Reyes JC, Rada-Iglesias A, de la Cruz X, and Martinez-Balbas MA (2018). Lineage specific transcription factors and epigenetic regulators mediate TGFbeta-dependent enhancer activation. Nucleic Acids Res10.1093/nar/gky093.
Irmak D, Fatima A, Gutierrez-Garcia R, Rinschen MM, Wagle P, Altmuller J, Arrigoni L, Hummel B, Klein C, Frese CK, Sawarkar R, Rada-Iglesias A, and Vilchez D (2018). Mechanism suppressing H3K9 trimethylation in pluripotent stem cells and its demise by polyQ-expanded huntingtin mutations. Hum Mol Genet 27, 4117-4134.
Rada-Iglesias A, Grosveld FG, and Papantonis A (2018). Forces driving the three-dimensional folding of eukaryotic genomes. Mol Syst Biol 14, e8214.
Cruz-Molina S, Respuela P, Tebartz C, Kolovos P, Nikolic M, Fueyo R, van Ijcken WFJ, Grosveld F, Frommolt P, Bazzi H, and Rada-Iglesias A (2017). PRC2 Facilitates the Regulatory Topology Required for Poised Enhancer Function during Pluripotent Stem Cell Differentiation. Cell Stem Cell 20, 689-705 e9.
Ludwig KU, Bohmer AC, Bowes J, Nikolic M, Ishorst N, Wyatt N, Hammond NL, Golz L, Thieme F, Barth S, Schuenke H, Klamt J, Spielmann M, Aldhorae K, Rojas-Martinez A, Nothen MM, Rada-Iglesias A, Dixon MJ, Knapp M, and Mangold E (2017). Imputation of orofacial clefting data identifies novel risk loci and sheds light on the genetic background of cleft lip +/- cleft palate and cleft palate only. Hum Mol Genet 26, 829-42.
Nikolic M, Papantonis A, and Rada-Iglesias A (2017). GARLIC: a bioinformatic toolkit for aetiologically connecting diseases and cell type-specific regulatory maps. Hum Mol Genet 26, 742-52.
Rada-Iglesias A (2017). Ready, Set...Poised!: Polycomb target genes are bound by poised RNA polymerase II throughout differentiation. Mol Syst Biol 13, 950.
Rehimi R, Bartusel M, Solinas F, Altmuller J, and Rada-Iglesias A (2017). Chromatin Immunoprecipitation (ChIP) Protocol for Low-abundance Embryonic Samples. J Vis Exp 10.3791/56186.
Information from this funding period will not be updated anymore. New research related information is available here.
Genomic context and mechanisms of transcriptional repression during early patterning of the mammalian embryo

Institute ofInstitute of Biomedicine and Biotechnology - University of Cantabria, ESomedicine and Biotechnology of Cantabria
CMMC - assoc. RG 18 (former CMMC JRG Leader)
show more…+49 221 478 96988
+49 221 478 97902
Institute ofInstitute of Biomedicine and Biotechnology - University of Cantabria, ESomedicine and Biotechnology of Cantabria
present address: Calle Albert Einstein 22
39011 Santander
http://zmmk-ari.uni-koeln.de/Rada-Iglesias_lab___Developmental_genomics/Home.html
CMMC Profile Page
Michaela Bartusel (doctoral student)
Tore Bleckwehl (doctoral student)
Tomas Pachano (doctoral student)
Rizwan Rehimi (postdoc)
Giuliano Crispatzu (postdoc)
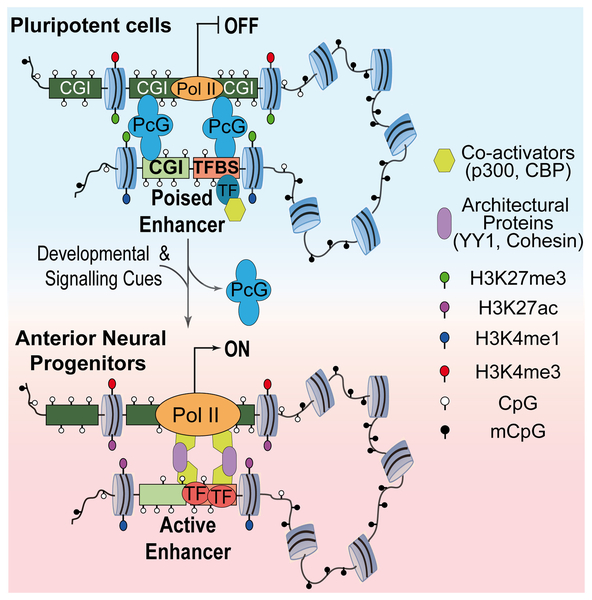
Current model of poised enhancer function during the differentiation of pluripotent cells into anterior neural progenitors.
Copyright ©
Prof. Dr. med. Thomas Benzing
Chair of the CMMC