Variations in the human FTO gene have been linked to obesity, altered connectivity and function of the dopaminergic neurocircuitry. We report that FTO in D2 medium spiny neurons (MSNs) of mice regulates the excitability of these cells in vitroand in vivo. Lack of FTO in D2 MSNs translates into increased locomotor activity to novelty in particular, without impairing the ability to control actions or affecting reward-driven and conditioned behaviour.
The fat mass and obesity-associated protein (FTO)is the first RNA demethylase identified, but the physiological role of FTO-dependent methyl-adenosine regulation remains only partially defined. In mice, ubiquitous inactivation of Fto reduces adiposity and protects from diet-induced obesity, and overexpression of FTO increases fat and body mass. However, whether FTO polymorphisms affect obesity development through altered regulation of FTO or other genes is not fully resolved. FTO polymorphisms are also associated with traits that are not restricted to overweight. Along these lines, Fto-deficient mice exhibit altered spontaneous and cocaine-induced locomotor activity, which is accompanied by impaired activation of the ventral tegmental area-substantia nigra (VTA-SN), and the striatum, brain areas critically involved in motor planning, decision-making, reward perception and judgment of time. Moreover, deletion of Fto in dopamine (DA) midbrain neurons impairs dopamine-2/3 (D2/3R) receptor-mediated inhibition of DA-signalling, resulting in enhanced sensitivity to the locomotor- and reward-stimulatory actions of cocaine.
Consistent with a role for FTO to regulate dopaminergic signalling, human FTO variants have been linked to changes in reward sensitivity, impulsivity, food reinforcement, preference for energy-dense food and augmented cortical activation in response to visual food cues. Collectively, all these data point toward a regulatory role for FTO in the integrated control of several behaviours governed by dopaminergic signalling. Nevertheless, while prior studies revealed that FTO controls D2/D3R-controlled auto-inhibition of DA neurons and that specific FTO genotypes alter midbrain responses to rewards, it is unclear whether FTO regulates the activity of basal ganglia neurons extending beyond the midbrain. Therefore, in the current study we investigated a potential role of FTO in controlling dopaminoceptive medium spiny neurons (MSNs).
To assess the potential physiological role of FTO through its action in neurons of the direct or indirect pathway, we generated mice lacking Fto selectively in D1 or D2 MSNs, i.e. Fto∆D1aor Fto∆D2mice. Given FTO’s role in regulating energy homeostasis, we assessed the metabolic phenotype of mice lacking FTO in D1 or D2 MSNs and found unaltered food intake, energy expenditure and body weight compared to control mice.
In light of the MSNs’ well-defined role in motor control, we also investigated diurnal activity patterns and found no alteration in circadian locomotor activity in both knockout lines in a domestic environment. To provoke ambulatory movement, we next examined the acute locomotor response to a novel environment. While Fto∆D1amice covered the same distance as their corresponding controls, Fto∆D2mice travelled substantially longer and were faster than wild-type littermates (Fig. 1). Further analyses revealed that Fto∆D2mice move faster and cover larger areas, have reduced stand, longer stride length, and higher swing speed. Combined, these results demonstrate that Fto∆D2mice exhibited hyperlocomotion acutely when exposed to a novel environment, whereas Fto∆D1amice show no motor changes.
To further investigate the hypothesis that the increase in activity was linked to novelty, we examined an array of behaviours during novel object recognition tests. While our results so far show that FTO’s actions in neurons harbouring D2Rs is involved in controlling explorative behaviour, they do not distinguish between pre- versus post-synaptic effects. Therefore, to differentiate between these two classes of D2R neurons and since FTO modulates D2R-like dependent control of DAT-neuron activity, we complemented the analysis with mice in which we ablated Fto in dopamine neurons (Fto∆DAT), i.e. neurons pre-synaptic to MSNs. Our data show that Fto∆D2mice had a higher preference for the novel object over the familiar, and to a larger extent than controls. Collectively, these results demonstrate that the behavior emerging from inactivating Ftoin D2 MSNs is induced upon exposure to novelty. These observations also indicate that the critical nexus underlying the observed behavior is post-synaptic D2 MSNs, as Fto∆DATmice - lacking FTO pre-synaptically - failed to replicate the same behavior as Fto∆D2mutants - lacking FTO pre- and post-synaptically.
In light of these results, we next examined the electrical properties of D2 MSNs ex vivoin order to gain insights into the consequences of removing Fto from D2 MSNs at the cellular level. We used genetically marked D2 MSNs by expressing tdTomato in D2R-expressing neurons in a Cre-dependent manner. Electrophysiological recordings revealed that the resting membrane potentials and the action potential (AP) thresholds for Fto∆D2MSNs were unaltered compared to FtowtMSNs. Furthermore, several AP parameters were unaltered by the lack of FTO in these neurons. However, when provoking APs with step and current injection pulses, pronounced changes between genotypes were observed in the voltage response, and in the current-voltage and current-frequency relationships of Fto∆D2and FtowtMSNs. Strikingly, less current was needed to depolarize the membrane and evoke an AP of Fto∆D2neurons compared to their wild-type counterparts, which is consistent with increased excitability (Fig. 2A).
To examine whether these differences in excitability would accompany a similar change in neuronal activity in vivo and to concurrently define D2 MSN activity dynamics during behaviour, we next performed fiber-photometry-assisted Ca2+imaging in awake mice based on viral- and Cre-induced expression of the calcium indicator GCaMP6s. We found that exposure to a novel environment led to a stronger increase in fluorescence intensity ratios in FtoKo-GCaMP6s+compared to FtoWt-GCaMP6s+mice, thus demonstrating augmented D2 MSN activation in the intact brain of mice lacking FTO in D2 MSNs upon novelty (Fig. 2B).
Ruud, J., et al., (2019). Fat mass and obesity-associated protein (FTO) regulates locomotor responses to novelty via D2R Medium Spiny Neurons, Cell Rep.27:3182–98
Cheng, Y., Luo, H., Izzo, F., Pickering, B.F., Nguyen, D., Myers, R., Schurer, A., Gourkanti, S., Bruning, J.C., Vu, L.P., Jaffrey, S.R., Landau, D.A., and Kharas, M.G. (2019). m(6)A RNA Methylation Maintains Hematopoietic Stem Cell Identity and Symmetric Commitment. Cell Rep 28, 1703-16 e6.
Ding, X., Willenborg, S., Bloch, W., Wickstrom, S.A., Wagle, P., Brodesser, S., Roers, A., Jais, A., Bruning, J.C., Hall, M.N., Ruegg, M.A., and Eming, S.A. (2019). Epidermal mTORC2 controls lipid synthesis and filaggrin processing in epidermal barrier formation. J Allergy Clin Immunol10.1016/j.jaci.2019.07.033.
Hammerschmidt, P., Ostkotte, D., Nolte, H., Gerl, M.J., Jais, A., Brunner, H.L., Sprenger, H.G., Awazawa, M., Nicholls, H.T., Turpin-Nolan, S.M., Langer, T., Kruger, M., Brugger, B., and Bruning, J.C. (2019). CerS6-Derived Sphingolipids Interact with Mff and Promote Mitochondrial Fragmentation in Obesity. Cell 177, 1536-52 e23.
Ruud, J., Alber, J., Tokarska, A., Engstrom Ruud, L., Nolte, H., Biglari, N., Lippert, R., Lautenschlager, A., Cieslak, P.E., Szumiec, L., Hess, M.E., Bronneke, H.S., Kruger, M., Nissbrandt, H., Korotkova, T., Silberberg, G., Rodriguez Parkitna, J., and Bruning, J.C. (2019). The Fat Mass and Obesity-Associated Protein (FTO) Regulates Locomotor Responses to Novelty via D2R Medium Spiny Neurons. Cell Rep 27, 3182-98 e9.
Turpin-Nolan, S.M., Hammerschmidt, P., Chen, W., Jais, A., Timper, K., Awazawa, M., Brodesser, S., and Bruning, J.C. (2019). CerS1-Derived C18:0 Ceramide in Skeletal Muscle Promotes Obesity-Induced Insulin Resistance. Cell Rep 26, 1-10 e7.
Wang, L., De Solis, A.J., Goffer, Y., Birkenbach, K.E., Engle, S.E., Tanis, R., Levenson, J.M., Li, X., Rausch, R., Purohit, M., Lee, J.Y., Tan, J., De Rosa, M.C., Doege, C.A., Aaron, H.L., Martins, G.J., Bruning, J.C., Egli, D., Costa, R., Berbari, N., Leibel, R.L., and Stratigopoulos, G. (2019). Ciliary gene RPGRIP1L is required for hypothalamic arcuate neuron development. JCI Insight 4.
Brandt C, Nolte H, Henschke S, Engstrom Ruud L, Awazawa M, Morgan DA, Gabel P, Sprenger HG, Hess ME, Gunther S, Langer T, Rahmouni K, Fenselau H, Kruger M, and Bruning JC (2018). Food Perception Primes Hepatic ER Homeostasis via Melanocortin-Dependent Control of mTOR Activation. Cell 175, 1321-1335 e1320.
Dafinger C, Rinschen MM, Borgal L, Ehrenberg C, Basten SG, Franke M, Hohne M, Rauh M, Gobel H, Bloch W, Wunderlich FT, Peters DJM, Tasche D, Mishra T, Habbig S, Dotsch J, Muller RU, Bruning JC, Persigehl T, Giles RH, Benzing T, Schermer B, and Liebau MC (2018). Targeted deletion of the AAA-ATPase Ruvbl1 in mice disrupts ciliary integrity and causes renal disease and hydrocephalus. Exp Mol Med 50, 75.
Dodd GT, Lee-Young RS, Bruning JC, and Tiganis T (2018). TCPTP Regulates Insulin Signalling in AgRP Neurons to Coordinate Glucose Metabolism with Feeding. Diabetes 10.2337/db17-1485.
Dodd GT, Michael NJ, Lee-Young RS, Mangiafico SP, Pryor JT, Munder AC, Simonds SE, Bruning JC, Zhang ZY, Cowley MA, Andrikopoulos S, Horvath TL, Spanswick D, and Tiganis T (2018). Insulin regulates POMC neuronal plasticity to control glucose metabolism. Elife 7.
Lohr H, Hess S, Pereira MMA, Reinoss P, Leibold S, Schenkel C, Wunderlich CM, Kloppenburg P, Bruning JC, and Hammerschmidt M (2018). Diet-Induced Growth Is Regulated via Acquired Leptin Resistance and Engages a Pomc-Somatostatin-Growth Hormone Circuit. Cell Rep 23, 1728-1741.
Quinones M, Al-Massadi O, Folgueira C, Bremser S, Gallego R, Torres-Leal L, Haddad-Tovolli R, Garcia-Caceres C, Hernandez-Bautista R, Lam BYH, Beiroa D, Sanchez-Rebordelo E, Senra A, Malagon JA, Valerio P, Fondevila MF, Ferno J, Malagon MM, Contreras R, Pfluger P, Bruning JC, Yeo G, Tschop M, Dieguez C, Lopez M, Claret M, Kloppenburg P, Sabio G, and Nogueiras R (2018). p53 in AgRP neurons is required for protection against diet-induced obesity via JNK1. Nat Commun 9, 3432.
Schuler R, Seebeck N, Osterhoff MA, Witte V, Floel A, Busjahn A, Jais A, Bruning JC, Frahnow T, Kabisch S, Pivovarova O, Hornemann S, Kruse M, and Pfeiffer AFH (2018). VEGF and GLUT1 are highly heritable, inversely correlated and affected by dietary fat intake: Consequences for cognitive fuction in humans. Mol Metab 11, 129-136.
Timper K, Paeger L, Sanchez-Lasheras C, Varela L, Jais A, Nolte H, Vogt MC, Hausen AC, Heilinger C, Evers N, Pospisilik JA, Penninger JM, Taylor EB, Horvath TL, Kloppenburg P, and Bruning JC (2018). Mild Impairment of Mitochondrial OXPHOS Promotes Fatty Acid Utilization in POMC Neurons and Improves Glucose Homeostasis in Obesity. Cell Rep 25, 383-397 e310.
Jiang H, and Bruning JC (2018). Melanin-Concentrating Hormone-Dependent Control of Feeding: When Volume Matters. Cell Metab 28, 7-8.
Awazawa M, Gabel P, Tsaousidou E, Nolte H, Kruger M, Schmitz J, Ackermann PJ, Brandt C, Altmuller J, Motameny S, Wunderlich FT, Kornfeld JW, Bluher M, and Bruning JC (2017). A microRNA screen reveals that elevated hepatic ectodysplasin A expression contributes to obesity-induced insulin resistance in skeletal muscle. Nat Med 23, 1466-73.
Dodd GT, Andrews ZB, Simonds SE, Michael NJ, DeVeer M, Bruning JC, Spanswick D, Cowley MA, and Tiganis T (2017). A Hypothalamic Phosphatase Switch Coordinates Energy Expenditure with Feeding. Cell Metab 26, 577.
Jais A, and Bruning JC (2017). Hypothalamic inflammation in obesity and metabolic disease. J Clin Invest 127, 24-32.
Paeger L, Pippow A, Hess S, Paehler M, Klein AC, Husch A, Pouzat C, Bruning JC, and Kloppenburg P (2017). Energy imbalance alters Ca(2+) handling and excitability of POMC neurons. Elife 6.
Ruud J, Steculorum SM, and Bruning JC (2017). Neuronal control of peripheral insulin sensitivity and glucose metabolism. Nat Commun 8, 15259.
Information from this funding period will not be updated anymore. New research related information is available here.
Role of prepronociceptin (PNOC)-expressing neurons in the development of obesity

Policlinic for Endocrinology, Diabetes and Preventive Medicine
CMMC - PI - C 03
Executive Board Member
+49 221 478 32123
Policlinic for Endocrinology, Diabetes and Preventive Medicine
Kerpener Str. 62
50937 Cologne
Johan Ruud (PostDoc)
Jens Alber (technician)
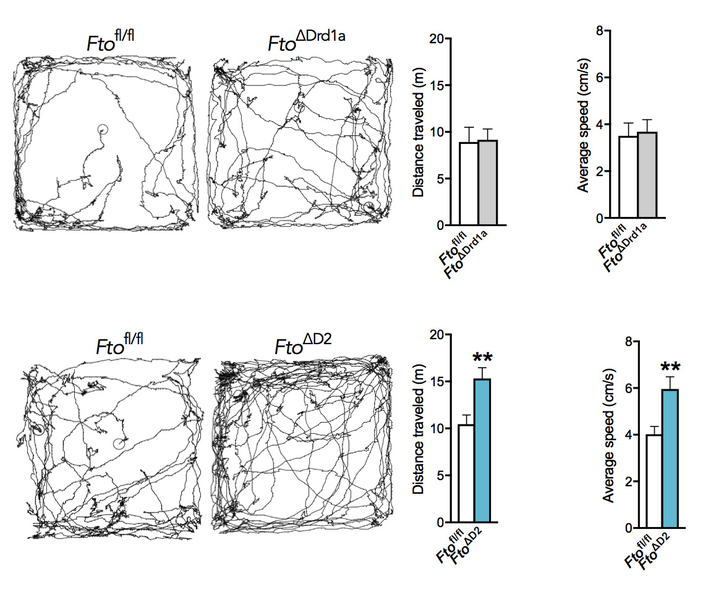
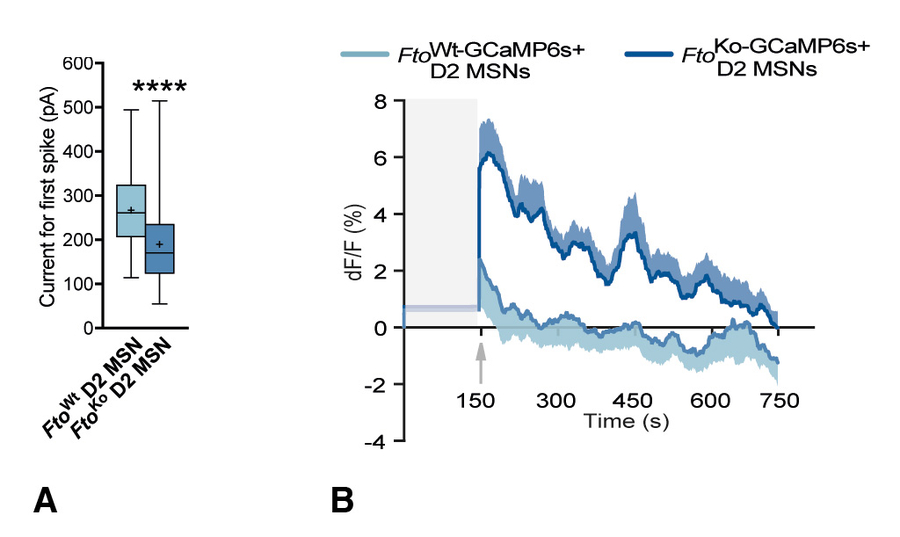
Fig. 2.FTO-deficient D2 MSNs display increased excitability ex vivoand in vivo.A) Box plot showing differences in minimum currents leading to first spike. FtoKoD2 MSNs were more excitable as significantly less current was needed to evoke discharge. B) GCaMP6s fluorescence-intensity before and after move to a novel enclosure in mice deficient in FTO in D2 MSNs and mice that had FTO intact in D2 MSNs. Shaded area represents time window for recordings of calcium-fluxes in home-cage environment (baseline for D2 MSN activity). Arrow marks the physical move to a novel enclosure.
Copyright ©
Prof. Dr. med. Thomas Benzing
Chair of the CMMC